
聽過「環烯醚萜」這個名字嗎?如果你告訴我你沒聽過,這一點也不意外,畢竟這個世界上的化合物太多太多了。不過,用來抗瘧疾的奎寧 (quinine),在它的合成路徑中,需要先合成環烯醚萜。也就是說,如果金雞納樹無法合成環烯醚萜,奎寧的合成就會有困難。
環烯醚萜(iridoid)是雙環單萜類次級代謝物,常見於菊類(asterids)植物。它的結構很特別,有一個五角環跟一個六角環。
說它有一個五角環跟一個六角環,可能有的讀者會想到吲哚(indole)。不過,吲哚是芳香性的「五角環 + 六角環」,由色胺酸、色素(如吲哚衍生物)、以及許多生物鹼(serotonin、tryptophan alkaloids)的核心骨架。
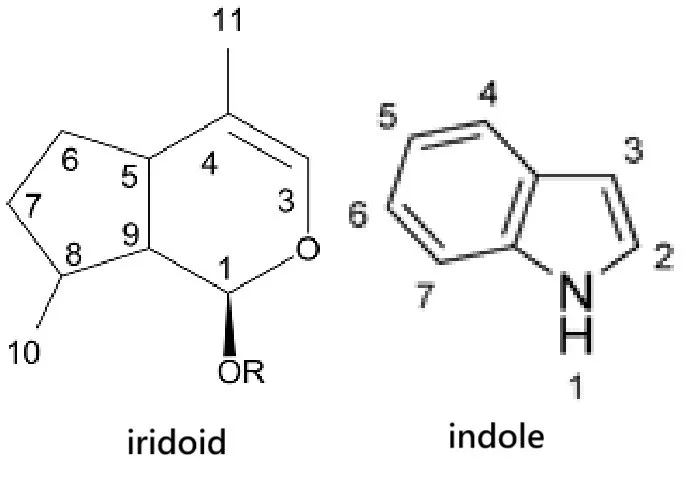
環烯醚萜(iridoid)與吲哚(indole)
而環烯醚萜則屬於非芳香性的「五角環 + 六角環」單萜衍生物,形成方式是由單萜中間產物經環化反應,得到雙環骨架。
植物合成環烯醚萜主要的目的是為了防禦。因此,除了大名鼎鼎的奎寧之外,當然還有更多的化合物屬於這個家族。有的具有揮發性,幫助植物吸引或驅逐昆蟲;有的則以糖苷形式存在,讓食草動物吃了「不舒服」。後來人類發現,這些化合物醫藥價值更大:環烯醚萜不僅本身有抗發炎活性,也是多種重要藥用生物鹼的前驅物,除了抗瘧疾的奎寧,還有抗癌的喜樹鹼 (camptothecin)、 、以及用來製造止吐藥吐根鹼(emetine)的原料。
也就是說,了解環烯醚萜的合成路徑,不僅僅是單純的基礎科學問題,更關係到人類如何利用植物化學來生產藥物。
過去的研究早就知道植物如何一步步合成環烯醚萜:從單萜前體 GPP(geranyl diphosphate)出發,經過一連串的轉換,最後由環烯醚萜合成酶(ISY 酶,Iridoid synthase) 生成不穩定的中間產物 8-oxocitronellyl enol。
這個中間產物非常不穩定,雖然會在試管裡會自己「摺起來」變成一個雙環結構 nepetalactol,但其自發性環化效率很低。雖然過去也因此,有些科學家以為,應該不需要額外的酵素,只靠自發反應就可以從8-oxocitronellyl enol到nepetalactol。
後來有人發現,在體外實驗裡,由中間產物形成的nepetalactol,大多都是 7S-cis-trans 型;但是在植物中,還同時存在著 7R-cis-cis 型的nepetalactol。也就是說,自發性的環化反應,即使我們可以忽略效率很低的問題,還是沒有辦法完全解釋在植物中看到的現象。
曾有許多研究團隊曾經試著把已知酵素(GES、G8H、8HGO、ISY)放到模式植物系統裡(例如菸草 Nicotiana benthamiana),卻始終無法完整重建 nepetalactol 的合成。
所有的這些發現都指向,環烯醚萜的合成還少了一個關鍵酵素。
另外,在唇形科的貓薄荷中有一個貓薄荷特有蛋白質(MLPL)能幫助生成 nepetalactol。MLPL(Major Latex Protein-Like) 是一類原本和「乳汁分泌」或「植物防禦」有關的蛋白質家族成員,並不是典型的代謝酶。它透過與 ISY 協同工作,把 8-oxogeranial 還原成非常不穩定的中間產物 8-oxocitronellyl enol。
他們發現,若單靠 ISY,反應效率極低,而且生成的多是錯誤的立體異構物;MLPL 能穩定中間產物,提供疏水環境讓其正確摺疊,並引導碳骨架在酵素活性位中以「正確角度」閉環,生成 7S-cis-trans nepetalactol。
因此,在有 MLPL 時,nepetalactol 生成量明顯上升,且產物立體化學一致。
相對的,若沒有 MLPL,反應不僅慢、產物也雜亂。
MLPL的例子啟發了研究團隊:或許在其他菊類植物,也有專門的酵素參與!
研究團隊以菊類植物土壇樹( Alangium salviifolium,山茱萸科,Cornales)與吐根(Carapichea ipecacuanha,Gentianales)為材料,使用了單細胞核 RNA 定序 (snRNA-seq),讓他們能看到在某些特定細胞類型裡,一整組環烯醚萜合成基因會同時被表現出來;從這些基因中,找出特定的候選基因放進菸草,和其他已知的環烯醚萜合成酶一起表現,看看是不是就是那個失落已久的酵素。
會選擇這兩種植物是因為,土壇樹屬於山茱萸目,是菊類(asterids)的基部分支之一。而吐根屬於 Gentianales(龍膽目),是另一個演化上相對遠離的支系。
由於這兩種植物在親緣關係上相距很遠,若在兩者都能找到同樣的基因,就能推論這個基因是整個菊類的祖先特徵,而不是個別族群才有的特殊演化。
另外是,吐根長期作為藥用(例如用於止吐的吐根鹼),它的環烯醚萜路徑活性很強,對這個題目來說是理想的研究材料。土壇樹雖然不是常見的藥用植物,但其基因體與代謝資料相對乾淨,容易建立對照。
結果他們發現其中的一個基因真的能推動反應完成,生成下游的loganic acid。他們把這個基因稱為環烯醚萜環化酶 (ICYC)。有趣的是,ICYC長得不像一般環化酶,反而更像「普通甲酯酶 (MES)」!通常我們在做基因搜尋時,多半是依賴序列相似性;因此過去都是找長得像環化酶的基因來測試,誰也不會想到要測試一個不像的酵素!
找到ICYC 之後,以它的序列進行搜尋後發現:原來在多數菊類植物中都有ICYC的存在,說明這是整個類群的「祖先特徵」。
特別的是,貓薄荷 (Nepeta)居然缺少 ICYC,因此自己演化出另一套獨立的環化機制,可說是一個「趨同演化」的好例子。
這個研究發現讓我想到,之前曾經跟學生提到的:科學是不斷嘗試著去接近真理的過程。但是,可能是因為技術限制,也可能是因為問錯問題,因此科學的發現都是「暫定」的,永遠都有可能被推翻。
過去為什麼找不到ICYC呢?因為過去科學家們只能用「相似性搜尋找出可能的標的基因 →逐一純化酵素或選殖酵素基因 → 加進試管或轉入模式植物 → 看有沒有產物出現」的方法。
但是在這裡,由於中間產物會進行自發性環化,這個「假象」讓人以為已經全員到齊;再加上ICYC長得不像環化酶,誰也不會想到它居然能催化環化反應。
因此,ICYC就這樣保持神秘了好多年。
這個發現提醒我們:
技術會改變我們能問的問題。沒有單細胞定序,我們可能永遠抓不到對的候選基因。
另外,問題如何設定,會影響能不能得到答案。由於過去一直認定ICYC應該要長得像一個「環化酶」(這是合理的假設),使得歷年來各研究團隊都是「過家門而不入」,始終與它失之交臂。
透過新技術的發展、問對問題,我們終於知道環烯醚萜的合成還需要再多一個酵素:環烯醚萜環化酶。
參考文獻:
Colinas, M., Tymen, C., Wood, J.C. et al. Discovery of iridoid cyclase completes the iridoid pathway in asterids. Nat. Plants (2025). https://doi.org/10.1038/s41477-025-02122-6


























