植物如何看見光?這個大哉問,應該是從達爾文看到窗邊的植物會朝著光源彎曲生長...或者,從更早的年代,就有人對為何植物可以看見光感到好奇了。
達爾文在1881年分享了他對植物向光性的研究,不過光敏素(phytochrome)的研究,大概還要更晚些。在Linda Sage的「Pigment of Imagination」一書中,有非常完整的敘述。
植物之所以能看見光,是因為有光敏素。光敏素就像植物的眼睛,讓植物可以看到光的顏色以及光的強弱。當然,從某種意義上來說,我們也可以說植物有點色盲,因為植物只會看它需要的光的顏色對不對,以及強度夠不夠強。所以,植物的光敏素只負責看紅光與紅外光,另外還有隱花色素(cryptochrome)與向光素(phototropin)是負責看藍光,而UVR8則負責看短波紫外光。
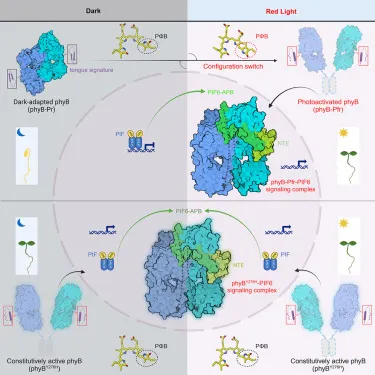
在找到光敏素(phytochrome)之後,科學家們也很快就發現,光敏素會因為光的照射而轉換構形(conformation),而這個構形的轉換,是因為光敏素中間的色素分子PΦB(phytochromobilin)在照光之後,構形產生變化-它的C15=C16雙鍵從順式(cis)構型轉變為反式(trans)構型,或者可以說,整個分子從「彎的」變成「直的」開始。
因為PΦB被包在光敏素分子的裡面,所以當PΦB「截彎取直」以後,與光敏素之間的互動就會開始產生變化。但是,這中間的詳細變化,以及發生變化後如何產生後續的效應,過去一直不是很清楚。
最近,有研究團隊以突變株的阿拉伯芥光敏素B與光敏素互動分子PIF6來進行冷凍電子顯微鏡,觀察到底光敏素看到光之後,會發生什麼事。
為什麼挑選光敏素B,是因為光敏素B是所謂的「主要」光敏素。在阿拉伯芥裡,光敏素A與光敏素B,負責主要的光感應工作,而光敏素B比光敏素A的重要性更高。缺少光敏素B的阿拉伯芥,在一般生長狀況下葉片呈現淡綠色且狹長,莖細細長長,節間大幅延長,開花時間大為提前,就是一副「我看不到光快不行了」的樣子。
為什麼要使用突變的光敏素B呢?因為野生型的光敏素B不會全部都停留在一個構形,就算給它照射高量的紅光或紅外光,也還會有大約3%的光敏素是另一個構形。為了觀察方便,研究團隊選了永遠保持活化的光敏素B突變種Y276H來進行研究。
當紅光照射到光敏素B的PΦB時,PΦB中的C15=C16雙鍵從順式(cis)構型轉變為反式(trans)構型(也就是整個分子從「彎的」變成「直的」),使得D吡咯環翻轉180°,這個變化使得位於光敏素B前端(N端)的PHY結構域發生了明顯的旋轉,導致PHY結構域的重新排列,並影響了其與GAF和nPAS等其他結構域的相互作用,從而改變了整個光敏素的結構。
其中有一個明顯的變化是PHY-tongue的出現。這個區域本來是β-片層結構(Pr狀態),而在紅光照射後會轉變為α-螺旋結構(Pfr狀態)。這個轉變是光敏素B活化過程中的關鍵步驟,因為它不僅影響了光敏素B的結構穩定性,還促進了它與光敏素互動因子(PIFs)的結合,從而啟動下游的信息傳導路徑。
另外,PHY結構域的變化會使得NTE結構域(N-terminal extension)變得更加靈活,並促進了NTE與光敏素互動因子(PIFs)的結合。這一結合不僅穩定了NTE的結構,還促進了光敏素B的活化,從而啟動了信號傳導路徑。
光敏素B在受到紅光照射後由Pr轉為Pfr,接著就會開始與許多不同的分子互動。為了了解光敏素如何與其它的分子互動,研究團隊選擇了PIF6來當代表,看看接下來會發生什麼事。
他們發現,PIF6與光敏素B的複合物是由兩個光敏素B和一個PIF6組成的。它們的結合主要涉及光敏素B的N端延伸域(N-terminal extension, NTE)以及光敏素特有的PHY域中的GAF模組。具體來說,PIF6的N端的第15-37胺基酸與光敏素B的NTE-nPAS-GAF模組結合,而PIF6的C端的第39-60胺基酸則被夾在GAF模組之間。這一結合不僅促進了PIF6的識別,還導致NTE的結構重塑,從而增強了光敏素B的穩定性。
當PIF6與光敏素B-Pfr結合後,這一複合物會進一步調控下游基因的表現,影響植物的生長和發育,例如促進種子發芽、葉片展開和花期調控等。
所以,透過研究光敏素B的結構,我們對於植物如何看見光更有了解,也希望從這些研究成果,可以更進一步了解光敏素如何將信息傳到其它下游的成員,讓植物生長發育!
參考文獻:
Wang et al., Light-induced remodeling of phytochrome B enables signal transduction by phytochromeinteracting factor, Cell (2024), https://doi.org/10.1016/j.cell.2024.09.005























