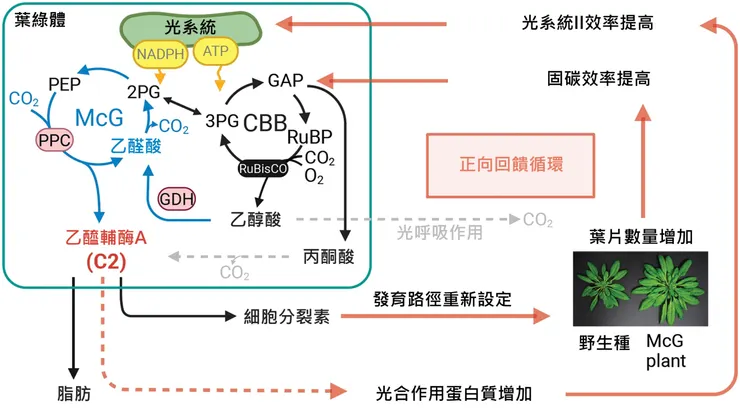
在植物細胞裡,乙醯輔酶A (acetyl-CoA) 是許多代謝途徑的起點,包括脂肪酸合成、植物激素(細胞激素、吉貝素、油菜素內酯等)的生合成,甚至還會影響蛋白質乙醯化訊號。但問題是:
葉綠體內的乙醯輔酶A主要依靠丙酮酸脫羧產生,不僅會損失固定的碳,而且效率不佳,所以能供應的相當有限。
再加上RuBisCO 的「雙重性格」:它既能固定 CO₂(好事),也會氧化反應產生 glycolate(壞事)。後者接下來就會進入光呼吸途徑,不僅耗能,還會排放 CO₂,白白浪費了辛苦固定下來的碳。
如果植物能更有效率地把這些碳轉成有用的代謝物,特別是乙醯輔酶A,是否可以讓植物的產量提升?
臺灣中央研究院廖俊智院士(也是現任中研院院長)帶領的團隊設計了一條人工合成代謝路徑,稱為蘋果醯輔酶A甘油酸酯 (malyl-CoA-glycerate,McG) 循環。他們的設計是:
他們將棒狀桿菌屬(Corynebacterium)的磷酸烯醇丙酮酸羧化激酶 (phosphoenolpyruvate carboxylase,PPC) 植入植物中,讓植物可以用它來固碳。由於PPC 使用的是碳酸氫根 (HCO₃⁻),不會跟 RuBisCO 搶 CO₂,就不會產生惡性競爭。
透過導入棒狀桿菌屬的PPC,讓植物可以用 3-磷酸甘油酸 (3PG)加上碳酸氫根,可以額外多固定一個碳,提升效率。
另外,他們還導入了萊茵衣藻(Chlamydomonas reinhardtii)的乙醛酸還原酶(glycolate dehydrogenase,GDH),於是當RuBisCO「看錯人」,產生乙醇酸的時候,就能把光呼吸的副產物重新導向,避免碳浪費。
但是,如果你以為他們只導入了棒狀桿菌屬的PPC與萊茵衣藻的GDH,那你就錯囉!研究團隊總共導入了六個外源基因,除了上述的兩個,還有:
莢膜甲基球菌(Methylococcus capsulatus)的 MTK(將蘋果酸轉換為 malyl-CoA)、MCL(把 malyl-CoA 分解成乙醯輔酶A和乙醛酸);
貪銅鉤蟲菌(Cupriavidus necator)的 GCL(把兩個乙醛酸拿來合成 tartronic semialdehyde)
大腸桿菌(E. coli)的 TSR(把 tartronic semialdehyde 還原成甘油酸 glycerate,回補循環)
這麼一來,阿拉伯芥就可以把由PPC抓下來的二氧化碳或由RuBisCO產生的乙醇酸,導引到合成脂肪的路徑了~
不過,要完成這個任務(生成乙醯輔酶A並合成脂肪),其實阿拉伯芥有四個酵素(MDH、PGK、PGAM、ENO) 也要加進來共同運作。
如此一來,乙醛酸的生成便不再是一種「浪費」,而是被拿去合成 acetyl-CoA。
當然,生物體往往比我們想像的要複雜得多,精心設計了一切,放進植物裡卻不能盡如人意,也不是沒有發生過。
那麼,這次呢?
將這套 McG 循環導入阿拉伯芥後,研究團隊觀察到:
植株乾重比野生種多了 2–3 倍,葉片更多更大,種莢和種子數量增加。
偵測光合作用效率時發現,淨 CO₂ 固定速率幾乎翻倍,光系統 II 效率 (ϕPSII) 提升,葉綠體中的「葉綠餅 (grana)」數量增加。
有趣的是,細胞激素(促進分生組織與葉片形成)含量升高,脂質含量顯著上升,葉片甚至出現可見的脂滴。
光呼吸作用造成的損失也減少了。他們發現,植物體內的乙醇酸濃度下降,CO₂ 流失降低。
值得注意的是,當他們把這個植物放在高 CO₂ 環境下生長時,McG 的優勢縮小;這意味著植物產生脂肪的途徑主要是透過把光呼吸副產物轉成乙醯輔酶A,而不是透過PPC來進行的。
這個研究是由中央研究院團隊完成,並於 2025 年 9 月發表在《科學 Science》期刊。
這不僅是植物代謝工程上的重大突破,也展現了臺灣在合成生物學、植物科學與全球糧食能源挑戰上的實力。
總而言之,透過植入其他生物的基因,研究團隊把「光呼吸的浪費」轉化成「脂肪的原料」。
另外,他們還建立了一個雙循環的碳同化新模式,並且展現了合成生物學如何解決傳統光合作用的瓶頸。
這不僅僅是一個研究成果,還展示了如何從基礎代謝問題出發,透過合成設計,重新規劃植物的碳流,並展現跨學科團隊的創新能量。
參考文獻:
Lu, K.-J., Hsu, C.-W., Chien, W.-N., Peng, M.-H., Chou, Y.-W., Huang, P.-S., Yeh, K.-C., Wu, S.-H., & Liao, J. C. (2025). A synthetic malyl-CoA–glycerate cycle enhances acetyl-CoA supply, lipid accumulation, and growth in Arabidopsis. Science, 373(6579), eadp3528. https://doi.org/10.1126/science.adp3528

























